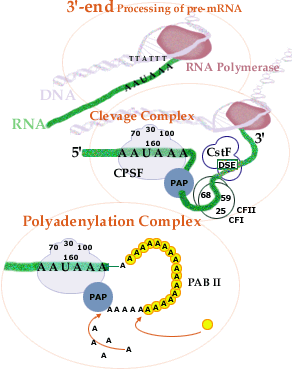
3'-end Cleavage and Polyadenylation.Virtually all mRNAs have a
number of
adenylate residues
at their 3'-end; the poly(A)-tail. These A's are not
encoded in the genes. Instead, the
pre-mRNA is cleaved
at a defined site and a poly(A) polymerase adds
150-200
adenylate residues.
The most important function of the poly(A)-tail is
during translation. The length of the poly(A)-tail
can be regulated in the cytoplasm. In some species,
for example, the egg cell stores mRNA in the
cytoplasm for later
use after fertilization. The stored mRNA has a short
poly(A)-tail. Activation of the mRNA for translation
includes lengthening of the poly(A)-tail. It has also
been suggested that the poly(A)-tail is involved in
determining how long an mRNA is present in the cell
before it is degraded. The 3'-cleavage and the initial part of the polyadenylation reaction take place at the active gene and polyadenylation is presumably completed as the pre-mRNA-protein complex is transported away from the gene. Polydenylation of pre-mRNA can be regulated in the cell in at least three different ways. First, there are alternative cleavage and polyadenylation sites for some genes. One example is during the development of B-lymphocytes. During this development, the synthesis of antibody molecules switches from membrane bound to secreted antibodies. The difference is due to a switch from one polyadenylation site to another. This results in a shorter antibody molecule, lacking a hydrophobic membrane-spanning part at the very end of the antibody molecule. Second, the very polyadenylation process can be modified. In one case, it has been shown that the protein product of an mRNA can bind to its own pre-mRNA and then inhibit polyadenylation. This results in degradation of the pre-mRNA and less production of the corresponding protein. Third, the expression of other genes can be regulated by changing the length of the poly(A)-tail in the cytoplasm. In some species, the oocyte stores mRNA in the cytoplasm for later use during the first cleavages after fertilization. These stored mRNAs have had their poly(A)-tail shortened in the cytoplasm. As the mRNAs are re-activated, their poly(A)-tails are extended by a cytoplasmic poly (A) polymerase. |